




서 론
고추장은 주원료로서 고춧가루와 전분질 (찹쌀이나 맵쌀), 발효에 관여하는 koji, 소금 등을 혼합하여 제조한 것으로, 독특한 맛을 가지면서도 기호성이 높은 우리나라의 고유한 복합 조미료이다(Kim & Choi, 2003). 일반적인 재래식 고 추장은 제조되는 지역이나 그 방법에 따라 염도의 차이가 크다.
전통식품의 CODEX 규격화 기술 지원 자료에 따르면, 총 13점의 전통 고추장 염도를 분석한 결과 약 7-9%까지 다양한 염도를 나타내었다고 보고된 바 있다(Park et al., 2001). 또한, 총 28점의 순창 고추장 민속마을의 전통 고 추장 제품의 염도는 약 5-10% 까지 나타난다고 보고된 바 도 있으며(Kim et al., 2006), 전라북도 지방의 전통 고추 장 15점의 성분을 분석한 결과에 의하면, 4.15-12.0%까지 편차가 매우 컸다(Cho et al., 1981).
염이 첨가되는 만큼, 장류를 많이 섭취할수록 식염의 섭 취량 또한 많아지게 되는데, 우리나라 국민이 평균적으로 섭취하는 식염의 약 73%는 장류에 의한 것이라고 한다 (Lim & Song, 2010). 식염의 주요한 성분인 나트륨을 다 량 섭취하게 되면 건강상의 문제를 일으킬 수 있기 때문에 세계보건기구(WHO)에서는 나트륨의 일일 최대 권고량을 2,000 mg으로 제한하고 있다. 이에 비해, 국민 1인당 1일 평균 나트륨 섭취량은 최대 권장량의 2.5-3배 정도에 육 박한다(Lee et al., 2007). 나트륨을 과잉 섭취하게 되면 고 혈압이나 심장질환, 혈관질환, 위암 등이 유발될 수 있기 때문에(Na et al., 1997), 질병에 대한 위험을 낮추기 위해 서는 맛과 보존성은 유지하면서도 염도를 최대한 낮추는 방안이 필요하다.
고추장의 품질은 단순히 원료 성분에 의한 것만이 아니 라, 고추장이 숙성되면서 생육하는 미생물의 발효에 의해 생성되는 대사 물질들도 많은 영향을 미친다. 따라서 본 연구에서는 염도를 달리한 전통 고추장이 발효기간에 따라 생육하는 미생물에 어떠한 차이를 보이는지에 대해 알아보 고자 하며, 더불어 고추장의 향미성분의 변화를 분석하여 염도를 달리한 고추장의 품질평가를 위한 자료를 얻고자 하였다.
재료 및 방법
식염이 첨가되지 않은 고추장은 순창장본가전통식품에서 구입하였으며, 염도를 달리하기 위해 신안군에서 제조한 갯벌천일염을 구입하여 염도에 맞게 첨가하였다. 식염을 첨가하지 않은 고추장의 구입 직후 염도는 3%였고, 여기 에 0.6 kg의 천일염을 첨가하여 6% 염도의 고추장을 제조 하였고, 1.4 kg 천일염을 첨가하여 10% 염도를 가지는 고 추장을 제조하였다. 또한, 2.4 kg 천일염을 첨가하여 15% 고추장을, 3.4 kg을 첨가하여 20% 고추장을 제조하여 숙성 기간별로 담금 직후(control), 숙성 1개월, 숙성 2개월, 숙 성 3개월, 숙성 4개월, 숙성 5개월, 숙성 10개월에 채취 하여 미생물 균총과 휘발성 향미를 분석하였다.
고추장 시료 10 g을 멸균 생리 식염수에 넣어 전체 부피 를 100 mL로 맞춘 뒤, 2시간 동안 균질화하여 시료 현탁액 을 제조하였다. 교반된 시료를 여과지(Whatman filter paper No. 2φ, 110mm)로 여과시켜 이물질을 제거한 후 여과액을 pH meter(model PH210, HANNA Instruments, Woonsocket, RI, USA)로 측정하였다.
염도별 숙성되는 고추장 시료에 존재하는 미생물 생균수 를 측정하기 위하여 십진 희석하여 101, 103, 105, 107, 109 의 희석액 500 μL를 nutrient agar(Difco Becton Dickinson, Cowley, Oxford, UK)배지에 도말하여 30°C에서 2일간 배 양한 후 형성된 집락을 계수하였다. 호염성 세균수의 측정 을 위해 십진 희석된 시료의 101, 103, 105, 107, 109의 희석액 500 μL를 HM agar(KCl 2.0 g, MgSO4·7H2O 1.0 g, CaCl2·7H2O 0.36 g, NaBr 0.23 g, NaHCO3 0.06 g, FeCl3 0.06 g, peptone 5.0 g, yeast extract 10.0 g, dextrose 1 g, agar 24.0 g, NaCl 100 g, distilled water 1.0 L) 배지에 도말하여 30°C에서 4일간 배양한 후 형성된 집락을 계수하였다. 효모의 균수 측정을 위해 101, 103, 105, 107, 109의 희석액 500 μL를 yeast mold agar(Difco Becton Dickinson, Cowley, Oxford, UK)에 20 μg/mL chloramphenicol(Sigma-Aldrich, St. Louis, MO, USA)을 첨가한 배지에 도말하여 25°C에서 4일간 배양 한 후 형성된 집락을 계수하였다.
균체 내 지방산 추출(Yang et al., 1993; Lee et al., 1996) 을 위해 해당 배지에 배양하였다. 호기성 세균 및 호염성 세균의 경우 trypticase soy agar(Difco Becton Dickinson, Cowley, Oxford, UK)에 30°C에서 24시간 배양, 효모의 경 우 YM 배지에 25°C에서 4일간 배양한 후 배양한 균체 40mg을 이용하여 지방산을 추출하였다. 시료의 분석 기기는 agilent technologies 6890 gas chromatography(Agilent Technologies model 6890N, Agilent, Palo Alto, CA, USA)를 사용하였고, separation column은 HP-1 column(cross-linked methyl siloxane, 30m long×0.320mm i.d., 0.25 μm film thickness, Agilent, Palo Alto, CA, USA)을 사용하였다. 표준 용액과의 비교에 의해 peak의 동정, retention time, peak의 면적, 면적 비율을 구하여 Sherlock MIS Software (Sherlock MIS ver. 6.2, Microbial ID, Inc. (MIDI), Newark, DE, USA)로 분석된 profile 비교를 통해 동정된 미생물명을 확인하였다.
고추장 향기 성분의 포집은 headspace SPME(Solid Phase Micro Extraction) 방법을 사용하였다. 향기성분의 추출을 위해 시료 고추장 3 g에 증류수 5 mL을 20 mL의 headspace glass vial에 넣어 준 후, 40°C의 water bath에서 30분 동안 vial을 평형상태로 만들어 주었다. SPME 1 cm 를 vial 안으로 주입하여 30분 동안 SPME fiber(50/30 μm DVB / Carboxen / PDMS, Agilent, Palo Alto, CA, USA)에 향기성분을 흡착시킨 후, SPME 3.5 cm를 10분 동안 GCMS( Gas Chromatography-Mass Spectrometer) injector에 넣 고 탈착시켜 향기성분을 분석하였다. Column은 CPWax- 52CB column(fused silica capillary, 50 m long×0.32 μm thickness, Agilent, Palo Alto, CA, USA)을 사용하였고, oven temp setting은 40°C에서 5분 동안 유지하였고 5°C/ min의 속도로 200°C까지 올려준 후, 200°C에서 20분간 유 지하였다. Carrier gas는 helium을 사용하였으며, 1.0 mL/ min의 속도로 흘려주었다. Injector 온도는 200°C, detector 온도는 250°C, ionization voltage는 70 eV, mass range는 33-350 m/z의 조건으로 진행하였다. 이 때 사용한 GC-MS는 GC(Agilent 7890A GC, Agilent, Palo Alto, CA, USA)에 detector인 MSD(Agilent 5975C, Agilent, Palo Alto, CA, USA)가 직접 연결된 mass spectrometer를 사용하였다.
결과 및 고찰
염도 별로 제조된 재래식 고추장의 숙성 기간 별 pH는 모든 염도에서 숙성 기간이 지날수록 pH의 감소 현상이 일어났지만, 3%와 6% 고추장의 pH가 훨씬 큰 폭으로 감 소하였다. pH가 급격하게 감소하는 현상은 6% 염도 고추 장보다 염도가 더 낮은 3% 염도 고추장에서 더욱 급격하 게 나타났다(Fig. 1).
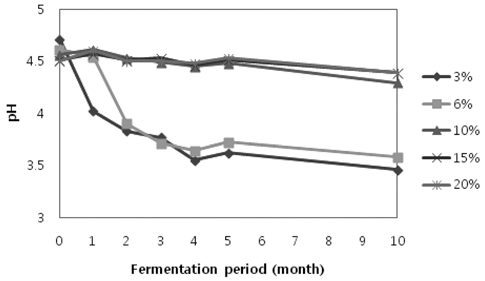
담금 직후에는 모든 시료의 pH가 pH 4.5 정도로 비슷한 수치를 보였다. 그러나 숙성이 진행될수록 염도에 따라서 pH 변화 양상이 달랐다. 담금 직후부터 숙성 2개월까지 3%와 6% 염도 고추장은 pH가 급격하게 감소하여 pH 4 이하로 떨어졌지만, 10%, 15%, 20% 염도 고추장은 큰 변 화 없이 pH 4.5 정도를 유지하였다. 숙성 3개월부터 5개 월까지는 3%와 6% 염도 고추장에서 꾸준하게 pH가 감소 하는 경향을 보이며 pH 3.5까지 감소하였고, 10%, 15%, 20% 염도 고추장은 큰 변화 없이 pH 4.5 정도를 유지하 였다. 염도 별로 담금 직후와 숙성 10개월의 pH 감소량 을 보면 3% 고추장은 숙성 10개월에 pH 1.25 감소하였 으며 6% 고추장에서는 pH 1.03 감소하였고, 10% 고추장 에서는 pH 0.27, 15% 고추장에서는 pH 0.12, 20% 고추장 에서는 pH 0.11 감소하였다. 염도가 7.8-9.9%인 전통 고추 장을 90일간 숙성시켰을 때 pH의 변화가 거의 없다고 보 고된 연구가 있으며(Cho et al., 1981), 재래식 고추장의 숙 성 기간 중 pH 변화를 나타낸 연구에서 소금 사용량이 적 은 고추장은 pH가 급격하게 감소한다고 보고된 바도 있다 (Kim et al., 1993). 이는 본 실험에서 10%, 15%, 20% 염 도의 고추장은 pH의 큰 변화가 없었으며, 염도가 낮은 3% 와 6%의 고추장에서는 pH가 급격하게 감소한다는 실험 결 과와 매우 유사했다. 염도가 낮은 고추장일수록 pH가 많이 감소하였으며 초기 pH 감소도 급격하게 일어났다. 식염이 많이 첨가 될수록 미생물의 생장이 저해가 되어 미생물 생 장 시 생성되는 유기산이 억제되어 숙성이 지나도 pH의 변화가 거의 일어나지 않는 것으로 보인다.
염도를 달리한 고추장의 호기성 세균과 호염성 세균, 효 모의 숙성 기간에 따른 생균수를 Fig. 2에 나타내었다. 호 기성 세균은 숙성 4개월까지는 모든 염도에서 큰 변화 없 이 7-7.5 log CFU/g의 수준으로 나타나고 있다. 그러나 숙 성 5개월이 지나면서 3%와 6% 염도의 고추장은 점차적 으로 감소해 각각 6.52, 6.40 log CFU/g까지 감소하였고, 숙성 10개월에 가장 염도가 낮은 3% 고추장은 6.20 log CFU/g까지 감소하였다. 6% 염도 고추장은 숙성 5개월에 6.40 log CFU/g까지 감소하였다가 숙성 10개월에는 이와 비슷한 6.47 log CFU/g로 나타났다. 반면에, 10, 15, 20% 염 도의 고추장은 처음의 수준을 계속 유지하였다. 염도를 달리 한 고추장의 일반세균의 생균수를 측정한 연구에 의하면, 5.9%, 6.8%, 8.5% 염도의 고추장 모두 숙성 초기부터 숙성 40일까지 생균수의 큰 변화 없이 6 log CFU/g 후반의 수 준을 유지하고 있는데(Lim & Song, 2010), 본 연구에서는 이보다 약간 더 높은 수준인 7 log CFU/g 정도로 나타나 기는 했지만 숙성 초기에 큰 변화가 없다는 결과는 동일했 다.
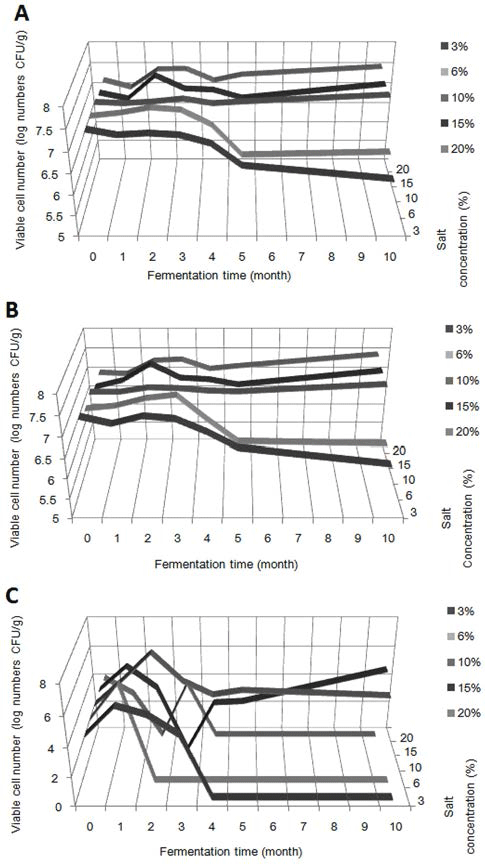
호염성 세균의 생균수 변화 추이는 호기성 세균과 유사 하였다. 숙성 3개월까지는 모든 염도에서 큰 변화 없이 7-7.5 log CFU/g의 수준을 유지하였다. 그러나 3%와 6% 고추장은 숙성 4개월부터 생균수가 점차적으로 감소되기 시작하여 3% 고추장은 숙성 4개월에 6.95 log CFU/g, 숙성 5개월에는 6.57 log CFU/g, 숙성 10개월에는 6.17 log CFU/ g까지 감소하였으며, 6% 고추장은 숙성 4개월에 6.87 log CFU/g, 숙성 5개월에 6.33 log CFU/g, 숙성 10개월에는 6.27 log CFU/g까지 감소하였다. 반면에, 10, 15, 20% 염도 고추장은 숙성 10개월까지 7-7.5 log CFU/g의 수준을 유지 하였다.
효모는 담금 직후 모든 염도의 시료에서 약 4.3 log CFU/g의 비슷한 생균수를 나타내었지만, 숙성이 지나면서 염도 별로 생균수의 변화 양상은 다르게 나타났다. 3%와 6% 고추장은 숙성 1개월에는 6 log CFU/g 정도로 생균수 가 증가하였다. 3% 염도 고추장은 숙성 2개월과 숙성 3개월에 각각 5 log CFU/g과 4 log CFU/g의 수치로 효모가 분리되었고, 그 이후에는 효모가 확인되지 않았다. 6% 염도 고추장에서는 숙성 1개월까지 6 log CFU/g로 효모가 분리 되었지만, 그 이후에는 효모가 확인되지 않았다. 재래식 고 추장의 숙성기간에 따라 효모 생균수가 증가했다가 숙성 90일경에 감소했다는 연구결과에서는 pH 저하가 원인이라 고 추정하고 있으며(Ahn, 1986), 딸기 퓨레를 첨가한 고추 장 역시 낮은 pH에 의해 숙성이 지날수록 효모 수가 감소 한다고 나타내었다(Kim & Lee, 2009). 본 연구에서도 3% 와 6% 염도의 고추장에서 pH가 저하가 급격하게 이루어 지면서 낮은 pH에 의해 효모가 생육하기 어려운 것으로 판단된다. 10%와 15% 염도의 고추장에서는 숙성 1개월에 두 가지 시료 모두 약 5 log CFU/g 정도의 생균수를 보여 주었으며, 15% 염도 고추장의 숙성 3개월 차에 효모가 분 리되지 않은 점을 제외하고는 두 가지 염도의 시료에서 모 두 숙성 10개월 차까지 꾸준히 효모가 분리되었다. 염도가 가장 높은 20% 고추장은 다른 염도와는 다르게 숙성 1개 월부터 생균수가 약 3 log CFU/g까지 감소하였으며, 숙성 4개월부터 효모가 분리되지 않았다. 10.5% 염도 고추장은 숙성 90일까지 효모 수가 꾸준히 증가했다는 연구 결과가 있으며(Kim & Lee, 2009), 소금의 종류를 달리한 10% 염 도 고추장의 경우에도 숙성 12주까지 꾸준히 효모가 분리 되었다는 보고가 있다(Kim et al., 2003). 숙성 초기 고추 장과 pH가 비슷한 수준인데 비하여 20% 염도 고추장은 숙성이 지나면서 효모가 분리되지 않았으며, 이는 pH 보다 는 높은 염도의 영향으로 효모가 생육하지 못한 것으로 사 료된다.
숙성이 지나면서 염도별 고추장 시료의 우점종의 변화 양상을 보기 위하여 해당 배지에서 임의로 5종씩 분리하 여 동정하였다. 3% 고추장에서 호기성 세균은 담금 직후 에 Peanibacillus macerans가 우점하고 있었고, 숙성 1개 월, 숙성 2개월까지는 Bacillus licheniformis, 숙성 3개월 에는 B. licheniformis와 더불어 B. amyloliquefaciens가 많 이 분리되었다. 숙성이 후기로 진행되면서 숙성 4개월, 숙 성 5개월, 숙성 10개월에는 B. subtilis가 3% 염도의 고추장 을 우점하였다. 6% 염도 고추장에서는 담금 직후와 숙성 1개월까지 B. subtilis가 지배균주였고, 숙성 2개월에는 B. pumilus, 숙성 3개월이 되면서 B. licheniformis가 우점하기 시작하여 숙성 5개월까지 분리되었다. 그러나 숙성 3개월 까지는 한 종만이 우점종인 반면에, 숙성 4개월, 5개월에 는 B. licheniformis 이외에 B. subtilis가 분리되었으며, 각각 B. megaterium와 B. atrophaeus 등 다양한 종이 분리되었다. 숙성 10개월까지 B. atrophaeus와 함께 B. subtilis는 많은 수 준으로 분리되었다. 10% 염도 고추장은 B. licheniformis와 B. subtilis 두 종만이 전 숙성기간 동안 우점종이었다. 숙성 1개월까지는 B. licheniformis와 B. subtilis 두 종 모두 우점 종이었지만, 숙성 2개월에는 B. licheniformis, 숙성 3개월 에는 B. subtilis가 각각 우점종이었고, 숙성 4개월, 5개월 에는 다시 두 가지 균주가 함께 분리되었며, 숙성 10개월 에는 B. subtilis만 우점종으로 나타났다. 또한 15% 염도 고추장은 10% 염도 고추장과 같이 B. licheniformis와 B. subtilis가 숙성기간 내내 우점종으로 확인되었다. 그러나 초 기 숙성 3개월까지는 B. subtilis만 높은 비율로 분리되었으 며, 숙성 4개월부터는 10% 염도 고추장과 동일한 우점종 을 보였다. 또한 20% 염도 고추장도 10%, 15% 염도 고추 장과 마찬가지로 주로 B. licheniformis와 B. subtilis가 숙성 기간 동안 우점종이었다. 숙성 3개월까지는 B. subtilis와 B. licheniformis가 교대로 우점종을 이루었고, 숙성 5개월에 Virgibacillus pantothenticus가 우점을 이룬 것을 제외하고 10%, 15% 염도 고추장과 숙성 기간 별 우점종을 이루는 균 주가 동일했다. 모든 염도에서 큰 비중으로 차지하고 있는 B. licheniformis와 B. subtilis는 고추장 발효에 중요한 역할을 하 는 것으로 사료된다. 염도가 낮은 3% 염도 고추장은 B. licheniformis와 B. subtilis 이외에 담금 직후와 숙성 3개월에 각각 P. macerans, B. amyloliquefaciens가 많이 분리되었고, 6% 염도 고추장 또한 숙성 2개월에는 B. pumilus, 숙성 4개월과 숙성 5개월에는 다양한 균주가 확인되었다. 10%, 15%, 20% 염도 고추장은 호기성 세균의 지배균주에 서 비슷한 경향을 보이는데, 주로 B. licheniformis 또는 B. subtilis가 모든 숙성기간 동안 가장 많이 분리되었다.
호염성 세균은 3% 염도 고추장의 담금 직후에는 호기성 세균에서 확인된 것과는 다르게 B. subtilis가 우점종을 이루 고 있으며, 숙성 1개월에는 B. subtilis 이외에도 B. atrophaeus, B. licheniformis, P. macerans 등의 다양한 균주가 우점종을 이루고 있다. 숙성 2개월부터 3개월까지는 다시 B. subtilis가 가장 많이 분리되었으며, 숙성 4개월에는 B. subtilis 이외에도 V. pantothenticus가 함께 우점종을 이루었 고, 숙성 5개월까지 많은 비중으로 확인되었다. 숙성 10개 월에는 B. subtilis와 함께 B. amyloliquefaciens, B. pumilus, P. larvae 등 다양한 종이 확인되었다. 6% 염도 고추장은 담금 직후에는 B. atrophaeus가 우점종을 이루었지만, 숙성 1개월부터 B. subtilis, B. pumilus, P. larvae가 함께 분리 되었으며, 숙성 2개월부터 숙성 10개월까지 꾸준히 B. subtilis가 6% 염도 고추장의 우점종이었다. 10% 염도 고 추장의 담금 직후에는 B. subtilis, 숙성 1개월에는 P. macerans가 우점종이었다. 숙성 2개월에는 B. subtilis와 B. licheniformis가 함께 큰 비중을 차지하였으며, 숙성 3개 월부터는 숙성 10개월까지는 6% 염도 고추장과 마찬가지 로 B. subtilis가 우점종이었다. 15% 염도 고추장은 10% 염 도 고추장과 마찬가지로 담금 직후에 B. subtilis가 우점종이 었고, 숙성 1개월에는 10% 염도 고추장과 같이 P. macerans가 많은 부분 차지하고 있었지만, 이외에도 B. atrophaeus, B. pumilus, Oceanobacillus 속 등 다양하게 확 인되었다. 그리고 숙성 2개월부터 숙성 10개월까지는 숙 성 5개월 차를 제외하고 6%, 10% 염도 고추장과 같이 B. subtilis가 우점종이었다. 20% 염도 고추장도 담금 직후 에 B. atrophaeus와 B. pumilus가 우점종이었지만, 숙성 1개월부터는 계속 B. subtilis가 20% 염도 고추장의 발효에 큰 비중을 차지하고 있다. B. subtilis와 함께 숙성 2개월에 는 B. licheniformis, 그리고 숙성 5개월에는 B. atrophaeus 가 많이 분리되었다. 호기성 세균과 마찬가지로, 호염성 세 균의 또한 B. subtilis가 모든 염도에서 가장 큰 비중으로 확인된다. 그러나 호기성 세균에서는 B. licheniformis도 함 께 고추장 시료에서 많이 분리된 반면에, 호염성 세균에서 는 비교적 적게 분리되었다. 반면에 Peanibacillus 속이 많 은 부분에서 우점종으로 확인되었다. 그리고 3% 염도 고추 장은 숙성 4개월부터 다양한 균종들이 우점을 하는 반면에, 6%, 10%, 15%, 20% 염도 고추장은 숙성 2개월부터 주로 B. subtilis가 우점종으로 확인되었다.
효모의 우점종을 보면, 모든 염도에서 숙성기간 동안 다 양한 균종이 확인되었다. 가장 저염인 3% 염도 고추장에 서는 담금 직후에 다양한 종이 분리되었다. 분리된 균주는 대부분 Candida 속이었으며, C. famata, C. parapsilosis, C. valida, C. zeylanoides가 확인되었다. 이 외에도 Cryptococcus terreus가 분리되었다. 숙성 1개월이 지나면서 C. valida와 함께 C. diversa, Kluyveromyces lodderii가 확인되었으며, 숙성 2개월에는 K. marxianus, 숙성 3개월에는 C. valida 가 우점종이었다. 또한 6% 염도 고추장은 3% 염도 고추 장과 같이 담금 직후에 C. valida가 우점종이었으며, 숙성 1개월에는 Saccharomyces cerevisiae가 가장 많이 분리되었 다. 3%와 6% 염도 고추장은 각각 숙성 4개월, 숙성 2개월 부터 pH 감소와 함께 균주가 분리되지 않았다. 10% 염도 고추장에서는 6% 염도 고추장과 같이 담금 직후에 C. valida가 우점종이었으며, 숙성 1개월과 숙성 2개월에는 S. cerevisiae가 가장 많이 분리되었다. 숙성 3개월에는 C. famata가 우점종이었으며, 숙성 4개월에는 다시 S. cerevisiae 가 많이 분리되었다. 숙성 5개월에는 S. cerevisiae 이외에도 C. famata, Cryp. albidus, K. lodderii, Rhodotorula rubra 등 다 양한 종이 분리되었다. 숙성 10개월에는 다시 S. cerevisiae이 많이 분리되었다. 15% 염도 고추장은 3%, 6%, 10% 염도 고 추장과 마찬가지로 담금 직후에는 C. valida이 많이 분리되었 고, 이 외에 C. fragariorum이 많이 분리되었다. 숙성이 지나 면서 숙성 1개월에는 C. famata이 지배균주였고, 숙성 2개 월에는 C. famata, C. polymorpha, C. utilis, C. zeylanoides, R. rubra 등 다양한 종이 확인되었다. 숙성 3개월에는 효모 가 분리되지 않았는데 pH의 감소에 의해 효모의 분리 유 무가 규칙적이지 않았던 Kim et al(1993)의 연구 결과와 같았으며, 본 연구 결과 역시 pH에 의해 효모의 분리 유 무가 규칙적이지 않은 것으로 사료된다. 숙성 4개월에는 높은 염도에서도 생육이 가능한 S. cerevisiae가 다시 우점 종을 나타내었으며, 숙성 5개월에는 C. versatilis, 숙성 10개월에도 C. halophila가 우점종이었다. 숙성이 되면서 높은 염도에서 생육이 불가능한 효모는 생육하지 못하는 것으로 보인다. 20% 염도 고추장에서는 다른 염도들과 마 찬가지로 담금 직후에는 C. valida이 지배균주였고, 숙성 1개월이 되면서 K. lodderii이 가장 많이 분리되었며, 숙성 3개월에는 S. cerevisiae가 우점종이었다. 가장 염도가 높은 20% 염도 고추장에서는 숙성 초기인 담금 직후와 숙성 1개월, 숙성이 조금 지난 후인 숙성 3개월에서만 균주가 분리되었다. 숙성 후기가 되면서 높은 염도에 의해 효모의 생장이 어려워지는 것으로 사료된다. 모든 염도에서 효모는 Candida 속과 S. cerevisiae가 고추장의 발효에 가장 중요 하게 관여하는 것으로 판단된다. 모든 염도에서 담금 직후 에 C. valida가 우점종이었다. 3% 염도 고추장에서는 숙성 기간 동안 Candida 속이 주로 발효에 관여를 하였고, 이외 에 Cryp. Terreus와 K. lodderii이 우점종이었다. 5가지 염 도 중 유일하게 S. cerevisiae가 우점종으로 확인되지 않았 다. 6% 염도 고추장에서는 C. valida와 S. cerevisiae가 숙 성 초기에 고추장 발효에 관여하였으며, 이후에는 효모가 분리되지 않았다. 10% 고추장도 숙성 초기에는 C. valida와 S. cerevisiae가 우점종으로 확인되었고, 숙성이 지나면서 C. valida와 함께 Cryp. albidus, K. lodderii, R. rubra 등 다양 한 종이 확인되었다. 15% 염도 고추장에도 C. valida와 S. cerevisiae 이외에 R. rubra가 숙성 2개월에 확인되었다. 20% 염도 고추장에서도 다른 염도와 마찬가지로 C. valida 와 S. cerevisiae가 우점종으로 확인되었고, K. lodderii이 숙 성 1개월에 우점종으로 확인되었다. 효모는 모든 염도에서 숙성기간 동안 다양한 종이 확인되었다.
본 연구에서는 휘발성 향미를 분석하기 위하여 향미 분석 방법 중 관능검사법 보다 객관적인 방법인 기기 분석방법을 사용하였다. 기기 분석을 이용한 방법은 향기성분에 대해 검 출 유무와 그 수치를 확인할 수 있으며, 관능 검사법에 비해 향미성분의 객관적인 판단이 가능하다(Noh, 2005). 기기 분 석 방법 중에서도 휘발성 향기성분들을 기체 이동상을 통해 단일 성분들로 분리해 주는 gas chromatography(GC)를 이용 하였다. GC와 질량분석계(mass spectrometry, MS)를 통해 동 정된 결과에 근거하여 고추장의 염도에 따라 숙성 기간별로 향미 성분의 변화가 어떻게 일어나는지를 연구하였다.
염도를 다르게 하여 제조한 재래식 고추장의 숙성과정 중 발생하는 휘발성 향기성분을 GC와 GC-MS로 분석하여 동정한 결과를 Table 1-5에 나타내었다. 재래식으로 제조된 염도를 달리한 고추장의 숙성 과정에서 alcohol류가 12종, aldehyde류가 5종, ester류가 26종, ketone류가 6종, furan류 가 3종, pyrazine류가 1종, acid류가 3종, sulfur-containing compound류가 1종, alkane류가 5종, alkene류가 5종, 기타 3종 등 총 70종의 휘발성 향기성분이 동정되었다. 계수적인 측면으로 보았을 때, ester류의 종류가 가장 많고, 그 다음 이 alcohol류, ketone류, aldehyde류의 순서로 나타났다.
고추장의 주요한 향기성분 중 하나인 alcohol 화합물은 발효를 통해 제조되는 주류나 고추장의 향기물질에서 중요 한 성분으로 보고되었다(Choi et al., 1997a; Choi et al., 1997b; Lee & Choi, 1998). Alcohol 화합물의 향기성분 중 에서는 모든 염도의 고추장에서 ethanol이 가장 높은 면적 비율을 나타내었다. 고추장의 고소한 향이 나는데 기여하 는 ethanol은 효모의 대사산물로, 당을 분해하면서 생성되 는 향기물질이다. Ethanol은 숙성 3개월과 4개월에 최대 검출량을 나타내었다. 3%, 6%, 20% 염도 고추장에서도 10-20% 정도의 높은 수준으로 나타났지만, 10%와 15% 염도 고추장에서 20-30% 정도로 상대적으로 높게 검출되 었다. 이는 10%와 15%에서 발효에 관여하는 효모가 가장 많이 증식하여 고추장의 향미에 영향을 주는 것으로 사료 된다. 2-Methyl-1-propanol과 3-methyl-1-butanol 또한 고추 장의 고소한 향미에 기여를 하는 성분이다. 2-Methyl-1- propanol은 3%, 6%, 20% 염도 고추장에서 0-1%의 미비한 수준으로 검출된 반면, 10%, 15% 염도 고추장에서는 이보 다 높은 1-2% 정도의 수준으로 검출되었으며, 숙성 3개월, 4개월에 최고 수준을 나타냈다. 또한 3-methyl-1-butanol도 10%, 15% 염도 고추장은 숙성 중기인 3개월, 4개월에 최고 수준인 10% 정도의 면적비율로 나타났지만, 3%, 6%, 20% 염도 고추장에서는 숙성이 지나면서 상대적으로 적은 양이 검출되었다. 고추장 원료인 대두의 비린내에 관 여하는 향기성분으로 보고된 1-pentanol과 1-hexanol도 검출 되었다(Wolf, 1975). 1-Pentanol은 발효 초기에는 높은 생산 량을 보이고 있으나 이후 숙성 4개월부터 함량이 현저히 줄어들었다. 그러나 3% 염도 고추장은 숙성 10개월에 다 시 1.69%의 높은 수준으로 성분이 검출되었다. 1-Hexanol 은 오히려 염도가 높을수록 높은 수준으로 검출되었다. 3%, 6% 염도 고추장에서는 1% 이하의 수준으로 검출된 반면에, 10%, 15% 염도 고추장에서는 숙성이 지나면서 1% 이상의 수준을 보였고, 20% 염도 고추장에서는 2% 정도로 검출되 었다.
Aldehyde류는 대부분의 시료에서 숙성 초기에는 검출되 지 않았으며 숙성 3개월부터 검출되기 시작하였으나, 검 출된 양은 다른 성분들에 비해 상대적으로 적은 양이었다. Aldehyde류 중 benzaldehyde는 crush bitter almond 향을 내며 식품에 인공적으로 첨가할 시 좋은 향미를 낼 수 있 으며, 메주와 된장에서 검출되는 주요 향기 성분이다(Kim et al., 1992; Kim et al., 1997). 다른 염도의 고추장에서는 숙성 3개월 이후부터 검출되어 0.5% 이하의 수준으로 나 타난 반면에, 20% 염도 고추장에서는 숙성 1개월부터 1% 이상의 수준으로 검출되어 숙성 5개월에 최고 수치인 2.52%로 상대적으로 많은 양이 확인되었다.Table.2
과일향을 내는 ester류는 분석된 70종의 향기성분 중 총 26가지로 가장 많은 성분이 확인되었다. Ester류는 alcohol 보다 휘발성이 강하기 때문에 고추장의 향미에서 가장 중 요한 역할을 하는 것으로 알려져 있다. 연구된 고추장 향 기성분에서도 ethanol과 함께 주요한 고추장 향기성분으로 보고되고 있다(Kim & Oh, 1993; Choi et al., 1997b). Ester 류 중 과실향과 감귤류의 주요한 향기 성분인 ethyl acetate 는 모든 염도의 고추장에서 담금 직후에 20-40%를 육박하 며 가장 큰 비율로 검출되었다. 특히 3%, 6%, 10% 염도 고추장은 대부분의 숙성기간 동안 20-30% 수준을 나타내 며 고추장 향기의 주성분으로 확인되었다. 15%와 20% 염 도 고추장은 숙성 3개월부터 10% 내외의 상대적으로 적은 양이 검출되었다. 더불어 3%와 6% 염도 고추장에서는 바 나나의 주요한 성분인 3-methyl-1-butyl acetate가 ester류 중 두 번째로 큰 비율로 검출되었다. 숙성 1개월부터 증가하 여 숙성 10개월까지 10-20% 수준으로 나타났다. 반면에 10% 염도 고추장은 숙성 10개월에 19%의 수준을 보였으 며, 이전 숙성기간 동안은 0-3% 정도의 상대적으로 적은 양이 검출되었다. 또한, 15%와 20% 염도 고추장도 전 숙 성기간 동안 검출되지 않거나 2% 이하로 검출된 것으로 보아, 10%, 15%, 20% 염도 고추장의 향미에 3-methyl-1- butyl acetate는 큰 기여를 하지 않는 것으로 사료된다. 감 미성 과실향의 ethyl hexanoate와 파인애플향의 ethyl octanoate는 공시된 고추장에서 낮은 면적비율로 검출된 성 분이다(Choi et al., 1997a; Choi et al., 1997b). 본 연구에 서도 3%, 6% 염도 고추장에서는 1% 이하의 낮은 면적비 율로 나타나거나 검출되지 않았지만, 감미성 과실향의 ethyl hexanoate가 10%, 15%, 20% 염도 고추장에서는 숙 성 3개월부터 2-5% 정도의 상대적으로 높은 비율로 확인 되었다. 또한 벌꿀향의 2-phenylethyl acetate는 모든 염도 에서 담금 직후에 많은 양 검출되었으며 이후에는 0.5% 이하의 낮은 면적비율로 나타났다(Yuda, 1976). 담금 직후 에 검출된 2-phenylethyl acetate는 염도가 높아질수록 높은 면적비율을 나타내는 경향이었으며, 15% 염도 고추장에서 가장 높은 14%의 면적비율을 가졌다. 숙성 초기와 후기에 각각 2-phenylethyl acetate와 ethyl hexanoate의 면적비율이 높았던 10%, 15%, 20% 염도 고추장이 3%와 6%의 고추 장보다 감미도가 높을 것으로 사료된다.Table.3
미생물의 중간 대사물질인 ketone류는 대부분의 연구된 고추장 향기분석과 마찬가지로 낮은 면적비율로 확인되었 다. 숙성 초기에는 검출되지 않은 성분이 더 많았으며, 숙 성 후기에 비교적 높은 빈도로 ketone류의 향기성분이 검출 되었고 검출된 향기성분의 수는 염도가 높을수록 많았다. 미생물 생균수 측정 부분에서 염도가 낮을수록 호기성 미 생물이 감소한다는 것을 확인하였는데, 이로 인해 미생물 이 대사하며 생성되는 ketone 물질이 적어지는 것으로 사료 된다. 이 경향은 미생물의 중요한 대사 물질이면서 발효유의 주요한 휘발성 향기물질인 3-hydroxy-2-butanone(Acetoin)에 서 극단적으로 확인되었다(Kim & Ko, 1993; Sun et al., 2012; Ann et al., 2013) 이 성분은 3%와 6% 염도 고추장 에서는 숙성 1개월부터 분리되지 않았지만, 10% 염도 고 추장에서는 숙성 2개월에 3% 이상의 수준으로 검출되었 으며, 15%와 20% 염도 고추장에서는 숙성 1개월과 2개 월에 10% 이상의 수준으로 검출되었다.
Furan류는 vinyl furan 및 2-pentyl furan, furfural이 비교 적 낮은 면적비율로 확인되었으며, 숙성 초기보다는 숙성 후기에 높은 빈도수로 검출되었다. 공시된 고추장 향기성분 에서는 당류가 산화할 시 생성되는 2-pentyl furan과 furfural 이 낮은 비율로 나타났으며, 본 실험에서는 고추장의 염도 가 높을수록 검출된 수치가 높았다(Kim & Oh, 1993; Choi et al., 1999).
Pyrazine류 중에서는 tetramethyl pyrazine이 전 숙성기간 동안 검출되었다. 이 향기성분은 benzaldehyde와 함께 청 국장의 쿰쿰한 냄새에 관여하는 성분으로 본 실험의 고추 장에서도 두 성분이 함께 고추장의 쿰쿰한 냄새에 관여하 는 것으로 사료된다(Kim et al., 1997). 위의 benzaldehyde 와 같이 고추장의 염도가 높아질수록 tetramethyl pyrazine 의 면적비율도 증가하는 경향이었다.Table.4
Acid류는 통상적으로 식품의 pH를 낮추는데 기여하는 성분이다. Acid류 중에서 미생물의 산화발효의 생성물인 acetic acid는 3%와 6% 염도 고추장에서 숙성 5개월까지 면적비율이 증가해 최고 검출 수치인 각각 19.45%, 25.91% 로 나타냈다. 반면에, 10%, 15%, 20% 염도 고추장에서는 10% 이하의 면적비율을 보여주면서 대부분의 숙성기간에 1% 정도로 낮게 검출되었다. 염도가 높은 고추장에서는 미생물의 생리활성저하에 의해 초산발효가 상대적으로 적 게 일어나는 것으로 보인다. Acetic acid는 식초에 다량으 로 포함된 향기성분으로써, 적은 양 존재할 때 단 과일향 을 내는 화합물이지만, 그 양이 많을 경우 오히려 본연의 고추장 맛을 저하시킬 수 있다(Yoon, 1998; Kim, 2010). 공 시된 고추장의 향미성분을 보면 낮은 면적비율로 acetic acid가 검출되는 것으로 확인되었다(Choi et al., 1997a; Choi et al., 1997b). 많은 acetic acid가 생성된 3%와 6% 염도 고추장은 본연의 고추장 향미보다는 식초와 같이 신 맛이 강할 것으로 사료된다. 다른 미생물의 산화 생성물인 3-methylbutanoic acid도 검출되었는데, acetic acid와는 대 조적으로 염도가 높은 고추장일수록 더 많은 수준으로 검 출되었다. 그러나 20% 염도 고추장에서도 5% 미만의 면 적비율로 검출되었으며, acetic acid 만큼 신 맛에 큰 영향 을 미치지는 않을 것으로 판단된다.Table.5
마늘, 파 등에서 매운 향을 내는 함황 화합물인 diallyl disulfide는 발효 기간에 따라서는 큰 차이를 보이지 않지 만, 염 농도가 높을수록 면적비율이 커졌다. 염의 농도가 높을수록 매운 향의 정도는 더 강할 것으로 사료된다.
Alkane류에서 hexamethyl cyclotrisiloxane, decamethyl cyclopentasiloxane 등 5종이 대부분 숙성 3개월부터 낮은 비율로 검출되었다. Alkene류 또한 3,3'-thiobis-1-propene과 3,7-dimethyl-1,3,7-octatriene 등 5종의 성분이 낮은 면적비 율로 검출되었다. 일반적인 재래식 고추장의 향기성분에서 도 alkane류와 alkene류는 적은 면적비율로 검출되며, 고추 장의 향기성분에 큰 영향을 주지 않는 것으로 보인다(Kim & Oh, 1993; Choi et al., 1997b).
기타 성분으로 toluene과 p-xylene, styrene이 검출되었다. 풀 향이나 benzene향을 가진 toluene은 숙성기간에 따라 비 교적 높은 비율로 검출되었다. 3% 고추장에서는 다른 고추 장에 비해 낮은 면적비율을 보였고, 6%, 10, 20% 고추장에 서는 중간 정도의 비슷한 수준으로 나타났으며, 15% 염도 고추장에서 10% 이상의 최고 면적비율을 나타내었다.
본 연구에서 검출된 총 70가지의 향기성분 중 alcohol류 의 ethanol, ester류의 ethyl acetate과 3-methyl-1-butyl acetate, ketone류의 3-hydroxy-2-butanone, acid류의 acetic acid 등이 숙성기간 동안 10-40%까지 높은 면적비율을 차지했다. 이 중 3-hydroxy-2-butanone는 3%와 6% 염도 고추장에서는 담 금 직후에만 소량 검출되었으며 그 이후에는 검출되지 않았 다. 또한, acetic acid는 상대적으로 3%와 6% 염도 고추장에 서만 다량으로 검출되어 일반적인 고추장의 향미보다 신 맛 이 강할 것으로 보인다.
요 약
pH의 변화는 10%, 15%, 20% 염도 고추장에서는 숙성 기간이 지나면서 약간의 pH의 감소 현상이 일어나기는 했 지만, 수치에 큰 변화는 없었다. 그러나 염도가 낮은 3%와 6% 고추장은 급격한 pH의 감소를 보여주었다. 호기성 세 균수는 숙성기간 동안 큰 변화가 없었지만, pH의 감소 경 향과 비슷하게 3%와 6% 염도 고추장에서 생균수가 더 많 이 감소했다. 모든 염도에서 B. licheniformis와 B. subtilis 가 큰 비중으로 차지하였다. 그러나 염도가 낮은 3%와 6% 염도 고추장에서는 이외에도 다양한 균주가 확인되었 다. 호염성 세균수의 변화 추이는 호기성 세균의 생균수 변 화 추이와 비슷했다. 호염성 세균도 B. subtilis가 모든 염도 에서 전 숙성기간 동안 가장 큰 비중으로 확인되었다. 그러 나 호염성 세균에서는 B. licheniformis 대신 Peanibacillus 속 이 많은 부분에서 지배균주였다. 이외에도 3% 염도 고추장 에서는 숙성 기간 동안 다양한 균주가 확인되었다. 효모는 분리 유무가 숙성기간에 따라 다르게 나타났는데, 이는 pH 의 급격한 저하에 의해 낮은 pH에도 생육이 가능한 균주 만 숙성후기에 분리되는 것으로 판단된다. 20% 이상의 염 도에서는 pH보다 고염에 의한 영향으로 효모의 증식이 어 려운 것으로 보인다. 모든 염도에서 Candida 속과 S. cerevisiae가 가장 많이 분리되었으며, 이외에도 다양한 균종 이 확인되었다. 검출된 휘발성 향기성분은 ester류가 26종으 로 가장 많고, 그 다음 alcohol류가 12종, ketone류가 6종 등 총 70종이 검출되었다. 이 중 alcohol류의 ethanol, ester 류의 ethyl acetate와 3-methyl-1-butyl acetate, ketone류의 3- hydroxy-2-butanone, acid류의 acetic acid 등이 10-40% 정도 의 면적비율로 검출되면서 주요한 성분으로 나타났다. 효 모의 대사산물이면서 고추장의 고소한 향미를 내는 ethanol은 10%와 15%에서 상대적으로 높은 비율로 검출되 었고, 과실향에 기여하는 ethyl acetate는 3%, 6%, 10% 염 도 고추장에서 상대적으로 높게 나타났으며, 반대로 발효 유의 중요한 향기성분인 3-hydroxy-2-butanone은 염도가 높아질수록 높은 비율로 검출되었다. 바나나의 향기에 중 요한 3-methyl-1-butyl acetate와 초산발효에 중요한 기여를 하는 acetic acid는 3%와 6% 염도 고추장에서 매우 높은 수치로 검출되면서 고추장의 이취를 내는 것으로 보인다. 결론적으로, 일반적인 고추장의 미생물 균총과 휘발성 향 미가 달라지지 않게 제조하기 위한 고추장의 최저 염도는 10%가 가장 적절할 것으로 사료된다.