서 론
두릅(Aralia elata Seem)은 두릅나무과에 속하는 낙엽관 목으로, 예로부터 민간과 한방에서 당뇨병의 주요 증상인 갈증을 해소하거나, 또는 갈증의 대사적인 원인을 제거하 는데 효과가 있으며, 항산화 효과, 당뇨유발 흰쥐의 유해활 성산소 생성저해 효과, 혈당저하 효과 및 HbA1C 수치 등 Biomarker 개선 효과 등이 보고(Lee et al., 1988; Lee & Kim, 1993; Kim et al., 2004; Shin, 2006; Lee et al., 2009)된 바 있다. 이 외에도 백내장 발생 억제 효과(Chung et al., 2005), 백혈암세포(HL60)와 폐암세포(A549)의 세포 독성(Zhang et al., 2013) 및 항염증(Lee et al., 2009; Nguyen et al., 2011), 심장기능 개선 효과(Wang et al., 2014) 등이 보고된 바 있으며, 두릅에 함유된 대표적인 생 리활성 화합물로는 triterpenoid saponins (Sim et al., 2005; Suh et al., 2007; Nhiem et al., 2011; Zhang et al., 2012; Kuang et al., 2013; Ma et al., 2013; Zhang et al., 2013; Wang et al., 2015)으로 알려져 있다.
유방암은 선진국형 질환으로, 전 세계적으로 매년 약 100만 명 이상의 여성들이 발병하고, 그 중 400,000명 이 상의 사망자가 발생하고 있다(Kamangar et al., 2006). 우 리나라의 경우, 발병 비율이 서구국가 보다는 상당히 낮은 것으로 나타나고 있으나, 서구화된 생활습관으로 인해 발 생률이 지속적으로 증가(Son et al., 2006)하고 있다. 유방 암 등 항암치료에 사용되는 대다수의 약제들은 암세포 뿐 아니라 정상 세포에도 영향을 미쳐 다양한 형태의 부작용 을 초래(Kim et al., 2004)하므로, 상대적으로 부작용이 적 은 약제로 개발된 다수의 천연물 유래 항암제들은 비교적 안전할 뿐 아니라, 항암효과를 상승시키는 것(Gouaze et al., 2004; Liu et al., 2007; Hatcher et al., 2008)으로 알려 져 있으며, 이와 관련된 연구가 광범위하게 진행되고 있다.
본 연구에서는 오랜 기간 건강소재로 활용한 천연재료로 부터, 인체에 안전하고 부작용이 적으며 인체 암세포 증식 을 억제하는 소재 발굴(Ryu et al., 2013b; Guon & Chung, 2014; Ryu & Chung, 2015)을 목적으로, 북한산 두릅 추출 물을 이용하여 MDA-MB-231 인체 유방암세포 증식에 관 한 성장억제 효과를 관찰한 후, MDA-MB-231의 세포사멸 (apoptosis) 및 세포주기(cell cycles)를 확인하였다. 이어서, 핵 형태 변화 및 유전자 발현에 미치는 기전 규명을 통해, 인체 유방암세포주인 MDA-MB-231 성장을 억제하는 천연 소재로 두릅의 활용 가능성을 관찰하였으므로 이에 보고하 고자 한다.
재료 및 방법
실험에 사용된 배지 RPMI 1640과 fetal bovin serum (FBS), penicillin-streptomycin (PS), Trypsin-EDTA는 GIBCO BRL (Grand Island, NY, USA) 제품을 사용하였고, MTT (3-(4,5-dimethydimethyl-2-yl)-2,5-diphenyltetrazolium bromide)와 propidium iodide (PI)는 Sigma Aldrich (St. Louis, MO, USA)에서 구입하였다. 1차 antibody인 β-actin, Bcl-2, Bax, caspase-3, caspase-9과 2차 antibody인 antimouse IgG HRP-conjugated와 anti-rabbit IgG HRPconjugated antibody는 Santa Biotechnology Co. (Santa Cruz, CA, USA)에서 구입하였다. 그 밖에 실험에 사용된 모든 시약은 공인된 등급의 순수용매를 사용하였다.
건조된 북한산 두릅 시료 지상부(30 g)를 70% ethyl alcohol로 90°C water bath에서 1시간 동안 2회 추출하고, 추출액은 여과지(Whatman No. 1, Cambridge, UK)와 초원 심분리기(Sorvall, Dupont Instruments, Wilminton, DE, USA)를 사용하여 여과하였다. 여과된 추출액을 rotary vacuum evaporator (Eyela, Tokyo, Japan)로 감압 농축한 후 -80°C에서 동결건조(Operon, Kimpo, Korea) 시켰다. 건 조된 시료(수득률 7.132 g)를 마쇄한 후 3차 증류수에 완전 히 용해시켜 실험재료로 사용하였다(Ryu et al., 2013b).
실험에 사용한 인체 유방암 세포주 MDA-MB-231과 인 체 대장암 세포주인 HCT116은 한국 세포주 은행(Korea Cell Line Bank, Seoul, Korea)에서 분양받아 10% fetal bovine serum과 100 units/mL penicillin, 100 μg/mL streptomycin을 첨가한 DMEM 배지를 사용하여 37°C, 5% CO2 incubator에서 배양하였으며, 세포밀도가 높아지면 Trypsin- EDTA를 처리하여 계대 배양하면서 실험에 사용하였다.
두릅 시료를 농도별로 처리한 후, 처리한 시료 농도별로 MDA-MB-231과 HCT116 세포증식 정도를 MTT assay로 측정하였다(Carmichael et al., 1987). 즉, MDA-MB-231 및 HCT116 세포를 96 well plate에 2×104 cells/well의 밀도로 분주하여 24시간 배양한 후, 시료의 최종 농도가 0, 50, 75, 100 μg/mL 되도록 처리하여 72시간 동안 배양하였다. 배양 후 각 well에 MTT 시약을 2.0 mg/mL 농도로 50 μL 를 첨가하여 4시간 동안 더 배양시킨 후 배지를 제거하고 각각의 well에 DMSO 150 μL를 첨가하였다. 이후 생성된 불용성의 formazan 결정을 용해시켜 570 nm에서 ELISA reader(Bio-Rad, Hercules, CA, USA)로 흡광도(optical density; OD)를 측정하였다. 세포증식률은 북한산 두릅 시 료의 흡광도를 대조군의 흡광도에 대한 백분율로 나타내었 고, 실험은 실험군마다 각각 3 wells의 흡광도를 측정하여 평균값을 계산하였으며 3회 반복 실시하였다.
MDA-MB-231 세포를 6 well plate에 3×105 cells/cm2 밀 도로 분주하여 24시간 배양한 후에 북한산 두릅 추출물을 농도별로 처리하여 24시간 동안 배양하였다. 배지를 제거 하고 PBS로 2회 세포 단층을 세척한 후, 100% methyl alcohol로 실온에서 30분 동안 세포를 고정하였다. 이어서 다시 PBS로 2회 세척하고 Hoechst 33258 최종농도가 2.0 μg/mL이 되도록 PBS에 희석하여 세포에 첨가하고, 15분 동 안 어두운 곳에서 염색한 형광현미경(Olympus Optical Co., Tokyo, Japan)으로 세포를 관찰하였다(Ryu & Chung, 2011).
Cell cycle 분석은 사멸된 세포들이 분포해 있는 sub-G1 의 비율을 측정하기 위해 수행되었다(Nicoletti et al., 1991). 두릅 추출물을 처리한 MDA-MB-231 세포를 모아 PBS로 세척한 후, 70% ethyl alcohol로 4°C에서 30분 동 안 고정시켰다. 그 후 다시 PBS로 washing하고, 암실에서 RNase A (50 μg/mL)가 포함된 propidium iodide (PI, 50 μg/mL) 용액을 첨가하여 세포의 DNA와 반응할 수 있도록 하였다. 이어서 FACS용 tube에 옮긴 후 FACS Calibur flow cytometry (Becton Dickinson, Franklin Lakes, NJ, USA)를 통해 측정하였다.
MDA-MB-231 세포에 두릅 추출물을 처리하였을 때, 세 포가 사멸된 정도를 정량적으로 분석하기 위해 flow cytometry를 이용하여 실험하였다. 세포사멸이 유도되면 세 포막의 phosphatidyl serine (PS)은 내부로 향하고 있다가 세포 외부로 노출하게 되는데, PS에 affinity를 갖는 annexin V를 사용하여 세포의 비율을 측정하고 PI를 사용하여 necrosis가 된 세포를 측정하였다. 두릅 추출물을 처리한 MDA-MB-23 세포를 모아 PBS로 세척한 후, 1×binding buffer (Annexin V-FITC Kit, Bechman courter, Pasadena, CA, USA) 100 μL를 첨가하여 부유시켰다. 부유된 세포액 에 발색시약으로 5 μL Annexin V와 0.7 μL PI를 첨가한 후, 암실에서 15분 동안 방치시켜 Annexin V와 PI가 세포 의 구성요소와 반응할 수 있도록 하였다. 그 후 FACS용 tube에 옮긴 후, FACS Calibur flow cytometry (Becton Dickinson)를 통해 사멸된 세포수를 측정하였다(Ryu & Chung, 2011).
북한산 두릅 추출물을 MDA-MB-231에 처리하였을 때 세포 증식을 억제하고 세포사멸을 유도하는 효과를 mRNA 범위에서 관찰하고, 세포사멸 시 modulator로 작용 하는 Bcl-2, Bax, caspase-9 및 caspase-3의 발현이 변화하 는 것을 확인하기 위하여 RT-PCR 방법을 수행하였다(Ryu et al., 2013a). 총 RNA 분리를 위해 Trizol reagent 1 mL 를 첨가하여 균질화시킨 후 chloroform 200 μL를 첨가한 뒤 4°C에서 13,000 rpm으로 20분간 원심 분리하여 총 RNA를 추출하였다. 추출한 RNA는 isopropanol을 사용하여 원심 분 리하여 정제하였고, 정제한 총 RNA는 diethylpyrocarbonate (DEPC)-water에 용해시킨 후 RNA 농도를 측정하였다.
다음으로 cDNA를 합성하기 위해 RNA 500 ng을 10 mM dNTP mix, Random hexamers (50 ng/μL), reaction buffer (10× RT buffer, 25 mM MgCl2, 0.1 M DTT, RNaseOUTTM), superscriptTM II RT, RNase H와 혼합하여 반응시켰다. 합 성된 cDNA를 template로 하여 Bcl-2 sense, 5'-AGC TGC ACC TGA CGC CCT TCA-3'; antisense, 5'-AGC CAG GAG AAA TCA CAG AGG-3'; BAX sense, 5’-ATG GAC GGG TCC GGG GAG CAG-3’; antisense, 5’-CAG TTG AAG TTG CCG TCA GA-3’; Caspase-9 sense, 5’- ATG GAC GAA GCG GAT CGG CGG CTC C-3’; antisense, 5’-GCA CCA CTG GGG GTA AGG TTT TCT AG-3’; Caspase-3 sense, 5’-TTC AGA GGG GAT CGT TGT AG-3’; antisense, 5’-CAA GCT TGT CGG CAT ACT GT-3’; β-actin sense, 5'-CCT CTA TGC CAA CAC AGT GC-3', antisense, 5'-ATA CTC CTG CTT GCT GAT CC-3'의 primer를 첨가한 후에, PCR mix buffer (10× PCR buffer, 25 mM MgCl2, 10 mM dNTP mix, Taq DNA polymerase)를 넣어 RT-PCR을 수행하였다.
시료 처리에 의한 MDA-MB-231 세포 사멸 및 세포 증 식에 관여하는 특정 단백질의 발현을 western blotting analysis를 실시하여 확인하였다(Ryu et al., 2013a). MDAMB- 231 세포를 60 mm dish에 분주한 후, 농도별로 시료 를 처리하여 72시간 배양한 후에 샘플을 모았다. 분석용 시료는 차가운 PBS로 2회 세척하여 2,000 rpm에서 10분 동안 원심분리 후에 세포를 수거하였다. 농도별로 모아진 세포는 1.0 mM PMSF와 proteinase inhibitor를 첨가한 NP40 cell lysis buffer를 첨가하여 30분간 세포를 얼음에서 용해시킨 후, 4°C에서 13,000 rpm으로 30분간 원심분리 후 상층액을 모아 분석시료로 사용하였다. 단백질 정량은 Bradford (Bio-Rad) 시약으로 측정하였으며, SDS-PAGE gel에 동량의 단백질을 loading하여 전기영동으로 분리한 후 단백질을 nitrocellulose membrane으로 전이시켰다. 단 백질이 옮겨진 membrane은 5% skim milk로 실온에서 1 시간 동안 blocking한 후, 1차 antibody를 처리하여 4°C에 서 하루 밤 반응시킨 후, TTBS buffer로 10분간 3회 세척 하고, 2차 antibody를 처리하여 실온에서 1시간 반응시켰다. 반응이 끝난 후 다시 TTBS buffer로 10분간 3회 세척한 후 enhanced chemiluminescence detection system (Amersham Phamacia, Amersham, UK)를 사용하여 특정 단백질의 발 현을 분석하였다.
결과 및 고찰
북한산 두릅 추출물이 MDA-MB-231 인체 유방암세포 성장에 미치는 영향을 확인하였다. 두릅 추출물을 0, 10, 50, 75, 100 μg/mL 농도로 처리하고, 각각을 72시간 동안 배양시킨 후, MTT 시약을 첨가하여 살아있는 세포가 미토 콘드리아 탈수소 효소반응으로 형성된 보라색의 formazan 을 확인하고 이를 DMSO로 용해시켜 흡광도를 측정하였 다. 실험결과, 시료를 0, 10, 50, 75 및 100 μg/mL 농도로 처리하였을 때, MDA-MB-231 세포 성장이 0%, 88.1%, 62.8%, 40.2% 및 5.0%로 나타났으며, 이는 MDA-MB-231 세포가 처리된 시료 농도와 비례적으로 성장이 억제되는 것이 관찰되었다(Fig. 1A). 이는 두릅에서 분리된 화합물인 Aralin을 여러 암세포에 44시간 동안 처리했을 때 IC50 값 이 HeLa 세포에서는 0.08 ng/mL, T24 세포에서는 2.6 ng/ mL, MIA PaCa-2 세포의 경우 5.1 ng/mL, MKN45 세포에 서는 8.5 ng/mL, Hep G2 세포의 경우 8.8 ng/mL, HL-60 세포에서는 10 ng/mL, OVK18 세포에서는 21 ng/mL의 세 포증식 억제 효과(Makoto et al., 2003)를 나타낸 것과 유사 하게 두릅 추출물도 특정 암세포의 성장을 지연시키는 것으 로 추정할 수 있다. 또한 인체 대장암세포주인 HCT116도 시료 농도와 비례적으로 증식이 감소하였으나, 본 실험에 서는 세포 사멸효능이 상대적으로 높은 MDA-MB-231 세 포주를 다음 실험에 사용하였다.
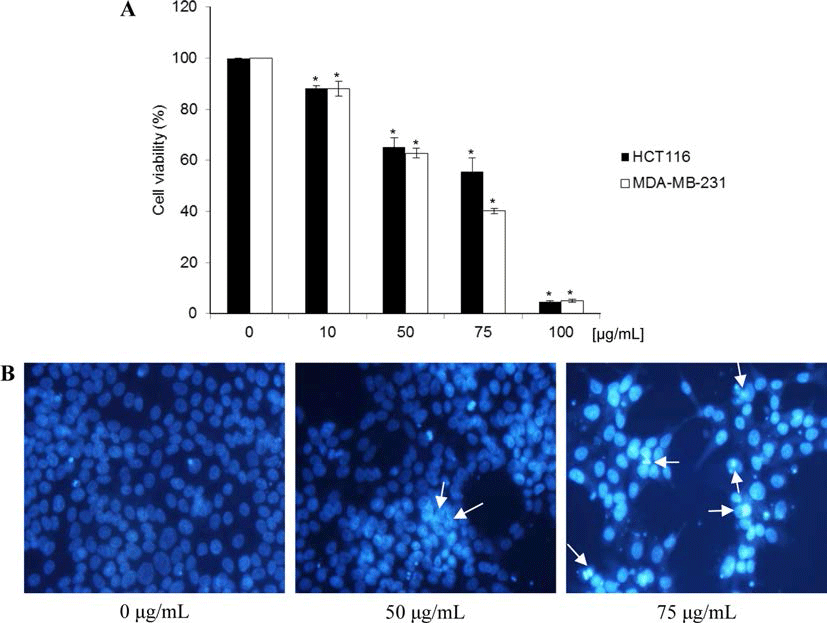
세포사멸(apoptosis)은 세포의 형태 변화 및 핵의 chroma tin 응축 등과 같은 일련의 변화로 확인될 수 있다(Gschwind & Huber 1995; Lizard et al., 1995). 두릅 추출물에 의해 MDA-MB-231 세포증식이 억제되는 것이 확인되었으므로, 세포증식 억제가 세포사멸과 관련이 있는 것인지를 확인하 기 위하여, Hoechst 33258 염색에 의한 핵의 형태 변화를 관찰하였다. 실험 결과, 두릅 추출물을 처리하지 않은 대조 군의 핵은 손상되지 않은 형태로 모두 균일하였으나, 두릅 추출물을 50 μg/mL 농도로 처리하였을 때, 약간의 핵이 응 축되는 현상이 확인되었고, 또한 세포 수가 감소된 것을 관찰할 수 있었다. 그러나 두릅 추출물을 100 μg/mL 농도 까지 증가시킨 경우, 바닥에서 떨어진 세포 수가 증가하였 고, 핵의 분절, chromatin 응축 및 형광 강도가 증가하는 등 세포사멸로 인한 apoptotic bodies가 눈에 띄게 관찰되 었다(Fig. 1B). 이러한 결과를 통해 두릅 추출물에 의해 MDA-MB-231 세포증식이 억제되는 것을 관찰할 수 있었 으며, 이는 위의 MTT assay 결과와 일치하는 것으로 확인 되었다.
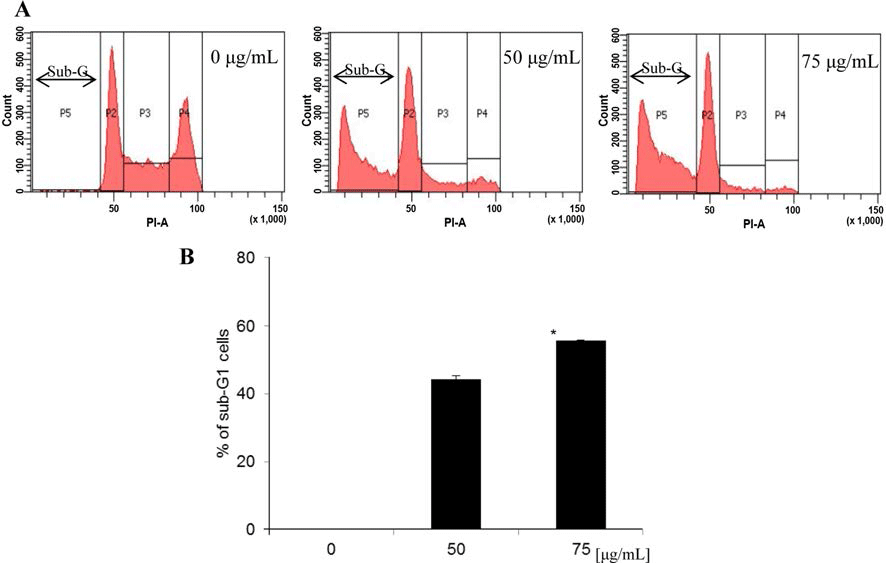
Cell cycle의 조절은 암세포의 성장과 분화 과정에 있어 중요한 역할을 하므로 항암 활성을 가진 물질을 대상으로 세포주기를 억제시키기 위한 연구들이 많이 진행되고 있으 며, 이때 sub-G1 증가는 사포사멸의 주요한 특징으로 알려 져 있다(Nicoletti et al., 1991). 이번 실험에서 두릅 추출물 이 MDA-MB-231 세포증식을 효과적으로 억제하였으므로, 세포증식 억제가 세포주기의 진행을 억제하여 일어난 것인 지 확인하기 위해 PI로 염색한 후 flow cytometry를 수행 하여 세포주기 변화를 관찰하였다. Fig. 2에 나타난 바와 같이, 시료를 처리하지 않은 대조군의 sub-G1은 0.04%인 반면, 두릅 추출물을 50 μg/mL과 75 μg/mL으로 처리한 경 우, sub-G1이 44.23% 및 55.64%로 나타나, 세포사멸의 특 징인 sub-G1의 수치가 대조군에 비해 증가됨을 확인하였다. S기에서는 시료 농도 0 μg/mL에서 28.30%, 50 μg/mL에서 11.19%, 75 μg/mL에서 6.79%로 감소하였고, G2/M기에서는 0 μg/mL에서 35.69%, 50 μg/mL에서 7.25%, 75 μg/mL에서 2.86%로 감소하여 세포주기가 억제됨을 관찰할 수 있었다. 이러한 결과는 MDA-MB-231 세포사멸 정도가 두릅 추출 물 농도와 비례적으로 발생되었고, 이는 시료 처리에 의한 세포 생존률 증가 및 세포 형태 변화와 일치하였다.
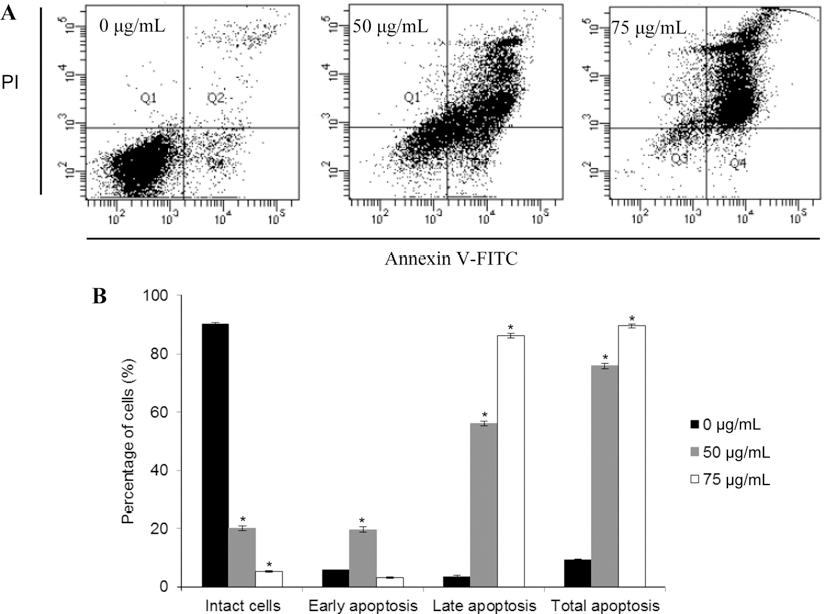
두릅 추출물에 의한 MDA-MB-231 세포증식 억제효과가 세포사멸(apoptosis)에 의한 것인지를 확인하기 위하여 Annexin V-FITC와 PI double staining을 사용하여 flow cytometric 분석을 통해 세포의 사멸정도를 정량적으로 분 석하였다. 분석 시 Annexin V-FITC-/PI는 대조군, Annexin V-FITC+/PI-는 early apoptosis, Annexin V-FITC+/PI+는 late apoptosis, Annexin V-FITC-/PI+는 necrosis를 의미한 다. Fig. 3A에 나타난 바와 같이, 시료를 처리하지 않은 대조군에 비해 두릅 추출물을 세포에 처리한 경우, 사멸된 세포의 수치가 유의적으로 증가되었다(p<0.05). 먼저, 대조 군의 early apoptotic 세포는 5.72%이었으나, 두릅 추출물 을 50 μg/mL으로 처리했을 때 19.72%로, 대조군에 비해 early apoptotic 세포가 증가하였다. 또한, late apoptotic 세 포의 경우 대조군에서 9.14%이었으나, 50 μg/mL와 75 μg/ mL으로 처리한 경우, 각각 56.16%와 86.45%로 late apoptotic 세포 수치도 시료 농도와 비례적으로 증가하였으 며, 특히 late apoptotic 세포가 early apoptotic 세포보다 더 많이 증가되는 것이 관찰되었다(Fig. 3B). 이상의 사실로, 두릅 추출물이 MDA-MB-231 세포의 apoptosis를 유도함 을 확인하였다.
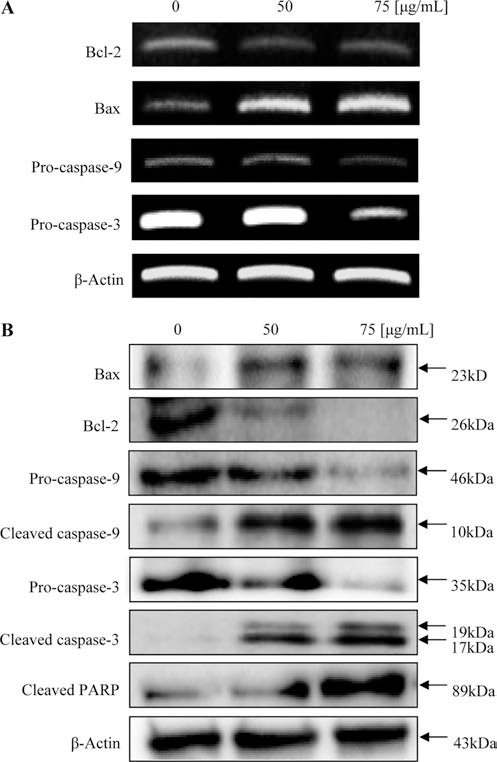
세포사멸과 관련된 신호체계(apoptosis signals)는 보편적 으로 extrinsic과 intrinsic의 두 가지 경로로 설명될 수 있 다. Intrisic 경로는 Bcl-2 family에 의해 조절되는데, Bcl-2 family는 각각의 기능과 아미노산 서열에 따라 크게 3가지 로 나뉜다(Wang, 2001). Chemotherapeutic 물질 등 외부 자극에 의해 세포가 사멸되는 것을 억제하는 Bcl-2 subfamily는 Bcl-2, Bcl-XL, Bcl-w가 있으며, 세포사멸을 유도하는 것으로는 Bax subfamily와 BH3 subfamily가 존 재하는데, Bax subfamily에는 Bax, Bak, Bok가 속하고, BH3 subfamily에는 Bad가 해당된다(Oltvai et al., 1993). 정상적인 상태에서는 미토콘드리아의 외벽에 존재하는 Bcl-2 subfamily가 Bax subfamily의 기능을 억제하며 서로 dimer 형태로 존재하여 균형을 유지하고 있다(Sattler et al., 1997). 그러나 외부자극으로 인해 이러한 균형이 깨지게 되 면, BH3 subfamily 단백질들이 활성화되어 Bcl-2 subfamily 기능이 억제되고, Bax subfamily들이 활성화되어 미토콘드 리아의 막 투과성이 증가되고, 미토콘드리아내에 존재하던 cytochrome c가 세포질로 방출된다(Cory & Adams, 2002). 따라서 Bcl-2와 Bax의 균형은 세포성장과 세포사멸에 중 요한 역할을 한다. 방출된 cytochrome c는 Apaf-1과 결합 하여 apoptosome을 형성하여 caspase endonuclease를 활성 화시켜 caspase-9과 caspase-3를 차례로 활성화시키고, 세포 사멸의 주요 인자인 PARP를 절단시키거나, 직접적으로 핵 의 응축과 DNA 절단에 영향을 주어 세포사멸을 유도한다 (Leung et al., 2007). 이러한 이론에 근거하여, 두릅 추출 물이 MDA-MB-231 세포사멸에 Bcl-2 family가 미치는 영 향을 확인하기 위하여 western blotting을 실시하였다. 실험 결과, Fig. 4A에 나타난 바와 같이, pro-apoptotic인 Bax의 발현이 시료 농도가 증가할수록 증가된 반면, antiapoptotic인 Bcl-2는 감소하였다. 또한, 불활성화 형태로 존 재하던 caspase-9과 caspase-3의 발현이 처리된 시료의 농 도에 따라 비례적으로 감소되었으며, 활성화된 cleaved caspase-9과 cleaved caspase-3의 발현은 두릅 추출물 처리 농도와 비례적으로 증가하는 것을 확인하였다. 또한 세포 사멸의 대표적인 표적인자인 PARP가 분할되는 것이 관찰 되었다. 따라서 두릅 처리에 의한 MDA-MB-231 세포사멸 은 Bcl-2/Bax 비율 변화에 의해 caspase가 활성화되는 것 으로 확인되었다.
요 약
본 연구는 인체 유래 유방암세포인 MDA-MB-231 세포 증식을 억제하고 세포사멸을 유도하는 천연소재 발굴을 목 적으로, 북한산 두릅 추출물을 MDA-MB-231 세포에 처리 하여 세포사멸 및 작용기전을 규명하였다. 실험 결과, 두릅 추출물 처리 농도가 증가할수록 세포증식이 감소하였고, 세포질의 응축과 핵이 분절되는 등 apoptotic bodies를 형 성하였다.
또한 유세포 분석기를 사용하여 MDA-MB-231의 세포주 기가 억제되었고, 세포사멸의 특징인 sub-G1 수치가 증가 되는 것이 관찰되었고, 두릅 추출물 농도가 증가함에 따라 apoptotic 세포가 유의적으로 증가하였으며, 특히 early apoptosis 보다 late apoptosis가 더 많이 진행됨을 확인할 수 있었다.
이를 바탕으로 MDA-MB-231 세포사멸과 관련된 유전자 를 확인하고자 RT-PCR과 western blotting을 수행한 결과, 세포사멸의 주요한 조절인자인 anti-apoptotic인 bcl-2 발현 이 두릅 추출물 처리 시 농도 의존적으로 감소되었고, 반 대로 Bax의 발현은 증가됨을 확인하였다. 이를 통해 불활 성화 형태로 존재한 pro-caspase-9과 pro-caspase-3의 발현 이 시료 농도가 증가할수록 감소되었고, 활성화된 형태인 cleaved caspase-9과 cleaved caspase-3의 발현은 두릅 추출 물 처리 농도에 의존적으로 증가하였다. 뿐만 아니라, 세포 사멸 과정의 주요 인자인 PARP 또한 시료 농도가 증가할 수록 절단 현상이 비례적으로 증가하였다.
이상의 실험 결과를 근거로, 북한산 두릅 지상부의 70% ethyl alcohol 추출물이 MDA-MB-231 인체 유방암 세포의 증식을 억제하는 효과가 있음을 확인하였고, 세포사멸과 관련된 활성기전을 규명하였다. 추후 암 예방/치료제로서 두릅을 활용하기 위해서는 활성 본체와 안정성 규명 및 임 상실험 등 추가 연구가 필요할 것으로 사료된다.